Research Report
Identification of QTL Involved in Low-temperature Tolerance at The Germination Stage by Recombination Inbred Lines in Rice 
2 Institute of Botany, Jiangsu Province and Chinese Academy of Sciences, Nanjing, 210014
 Author
Author  Correspondence author
Correspondence author
Rice Genomics and Genetics, 2020, Vol. 11, No. 3 doi: 10.5376/rgg.2020.11.0003
Received: 05 May, 2020 Accepted: 11 May, 2020 Published: 11 May, 2020
Zhang S.B., Zhang Y.H., Lin J., Chen H.Y., Wang Y.J., Zhu X.M., Song C.F., and Fang X.W., 2020, Identification of QTL involved in low-temperature tolerance at the germination stage by recombination inbred lines in rice, Rice Genomics and Genetics, 11(3): 1-6 (doi: 10.5376/rgg.2020.11.0003)
Low-temperature germination (LTG) resistance of direct-seeded rice (DSR) seeds can overcome the developmental delay caused by low-temperature stress and ensure the vigorous growth of seedlings. The LTG is complex quantitative trait controlled by QTLs and has low genetic force, which leads to low efficiency of the direct selection of the trait by breeders. The research on QTL mapping for LTG in rice is helpful to carry out molecular marker-assisted selection (MAS) and improve the efficiency of selection for LTG in DSR breeding. In this study, the LTG rate showed extremely significant differences between the high-quality japonica rice variety Nanjing 46 in Jiangsu and the local variety Zhaxima in Yunnan. The QTL mapping was carried out by a recombination inbred line population (RIL) derived from a cross between Zhaxima and the Nanjing46. Total three QTLs (qLTG-2, qLTG-4 and qLTG-7) involved in LTG were detected on chromosome 2, 4 and 7, respectively. They accounted for 7.69%, 8.75% and 22.93% of the total phenotypic variation (PV), respectively. Interestingly, there was no reported QTL involved in LTG in the candidate region of qLTG-2, which exhibited that the qLTG-2 was a novel QTL locus. The QTLs detected in this study will provide new genetic material and molecular markers for improving the low-temperature tolerance at germination stage in rice by molecular marker assisted selection breeding.
Rice (Oryza sativa L.) is one of the three major food crops in the world, providing a staple food for more than half of the world's population. Asia accounts for 90% of global rice consumption, and total rice demand of Asia continues to rise. China is the largest rice producer and consumer in the world, accounting for 30% of total world production and consumption (http://ricepedia.org). China has a history of rice cultivation for thousands of years. The traditional Chinese rice cultivation method is mainly based on artificial seedling raising and transplanting. This model contributes to the seedling formation and the effective use of nutrients (Farooq et al., 2011), and can effectively inhibit the growth of weeds (Singh et al., 2001; Olajumoke et al., 2016). With the rapid development of China's economy, the number of people engaged in agricultural labor has decreased sharply, resulting in a substantial increase in labor costs, and rice production investment has become more expensive and less profitable. Interestingly, DSR is a light, water-saving and efficient cultivation mode, which is also of great significance to alleviate labor shortage. DSR with strong seed germination ability under low temperature is one of the basic ability to ensure that they form strong seedlings. However, many high-yield and high-quality varieties show low germination rate under low-temperature conditions, which limits the development of DSR (Chen et al., 2006; Cao et al., 2018).
LTG of rice is a quantitative trait controlled by multiple genes. Chen et al. (2006) reported 2 QTLs for LTG ability, qLTG-3 and qLTG-10, by using 198 doubled haploid (DH) population derived from a cross between indica rice Zhenshan 97B and AAVOO2863. The qLTG-3 flanked by RM85 and RM514 on chromosome 3 and qLTG-10 flanked by RM596 and RM271 on chromosome 10 accounted for 12.6% and 12.9% of the total PV, respectively. The enhancing alleles came from Zhenshan 97B and AAVOO2863, respectively. Fujino et al.(2004) found 3 QTLs (qLTG-3-1, qLTG-3-2 and qLTG-4) correlated with LTG by using a RIL population derived from a cross between Italica Livorno and Hayamasari, which located on chromosome 3 and 4, and contributed to 35.0%, 17.4% and 5.5% of the total PV, respectively. According to the germination rate within 7 to 15 days, Jiang et al. (2006) revealed 9 QTLs (qLTG-3, qLTG-4, qLTG-5-1, qLTG-5-2, qLTG-5-3, qLTG-9, qLTG-10, qLTG-11-1 and qLTG-11-2) related to LTG in an F2 population derived from the cross USSR5×N22, which located on chromosome 3, 4, 5, 9, 10 and 12, respectively. The proportion of the PV explained by these loci was 4.16% and 31.23%, respectively. Ji et al. (2008) detected 11 QTLs using RIL population derived from Kinmaze (Japonica rice)/DV85 (Indica rice), of which qLTG-7 and qLTG-11 could be stably mapped in 3 different environments. Among them, the largest contribution to PV was up to 27.93%. Gong et al. (2009) revealed 4 QTLs (qLTG-1, qLTG-7, qLTG-9 and qLTG-11) of LTG in a backcross RIL derived from the cross Koshihikari/Kasalath//Koshihikari, which distributed on chromosome 1, 7, 9 and 11, respectively. The contribution to PV was from 6.72% to 12.78%. According to the germination rate within 7 to 15 days, Ji et al. (2009) found 6 QTLs (qGR-2, qGR-3, qGR-7, qGR-10, qGR-11 and qGR-12) related to LTG in an RIL population derived from the cross Asominori/IR24, which were on chromosome 2, 3, 7, 10, 11 and 12, respectively. Schlappi et al. (2017) uncovered 6 QTLs conferred LTG by 202 O. sativa accessions from the Rice Mini-Core collection, which lied on chromosome1, 6, 7, 10 and 12, respectively. According to LTG or stress index of LTG (LTGS) after 2, 4, and 6 days of sowing, Najeeb et al. (2020) reported that a total of 21 and 26 QTLs associated with LTG and LTGS by a BC1F7 population derived from the Weed Tolerant Rice-1 and Haoannong. These QTLs were located on all the chromosomes, except for chromosomes 12.
Although some QTLs related to LTG had been reported, the mechanism of LTG in rice was still unknown. Mapping, isolation, and functional analysis of novel QTLs related to LTG was helpful for elucidating the molecular mechanism of LTG. In this study, the QTLs related to LTG were identified by a RIL population consisting of 135 lines derived from a cross between the high-quality japonica rice variety Nanjing 46 in Jiangsu and the local variety Zhaxima in Yunnan, which would be benefit for fine mapping and isolation the QTLs and be helpful for providing theoretical basis for breeding utilization.
1 Results and Analysis
1.1 Analysis of the germination rate of parents and the RIL population
After germination at 15°C for 10 days, the germination rates of Zhaxima and Nanjing46 were investigated. The results showed that the germination rate of Zhaxima was 25.14%, while that of Nanjing46 was 84.99%. The difference between them reached a statistically significant difference (p<0.01) (Figure 1). The germination rate in the RIL population derived from a cross between Zhaxima and Nanjing46 showed a continuous distribution, which ranged from 0.00% to 100%. The frequency distribution diagram of RIL population exhibited skew distribution (Figure 2).
 Figure 1 Phenotype of Zhaxima and Nanjing46 under low-temperature conditions Note: A: Phenotype of Zhaxima (Left) and Nanjing46 (Right) for 10 d under low-temperature conditions; B: Rate of germination of Zhaxima and Nanjing46 for 10 d under low-temperature conditions |
 Figure 2 Frequency distribution of LTG rate in Zhaxima/Nanjing46 RIL population |
1.2 QTL analysis of LTG
In the previous study, a molecular marker linkage map of the RIL population from Zhaxima×Nanjing46 was obtained by 202 pairs of simple sequence repeat (SSR) primers, which evenly distributed on 12 chromosomes of rice, and exhibited polymorphic differences between Zhaxima and Nanjing46. The total length of the linkage map was 1 437.3 cM, and the average distance of the linkage map was 8.1 cM, which could meet the requirements of QTL mapping (Fang et al., 2017; Zhang et al., 2018). According to the rate of LTG in the RIL population of 135 lines, the location and genetic effect of QTL controlling LTG were analyzed by QTL Ici-Mapping 4.0. A total of 3 QTLs QTL (qLTG-2, qLTG-4 and qLTG-7) related to LTG were detected, which located on chromosome 2, 4 and 7, respectively. These QTLs explained 39.37% of the PV for LTG (Figure 3; Table 1). Among them, the highest PV was explained by qLTG-7 with a LOD score of 8.02, which explained 22.93% of the PV for LTG. These showed that the qLTG-7 was a QTL with a high effect value. The qLTG-7 had negative additive effect, indicating the enhanced LTG allele was from Nanjing46. The LOD values of the other two QTLs were 3.00 and 3.37, which could explain the PV of 7.69% and 8.75%, respectively. The two QTLs had positive additive effect, indicating the enhanced LTG alleles were from Zhaxima.
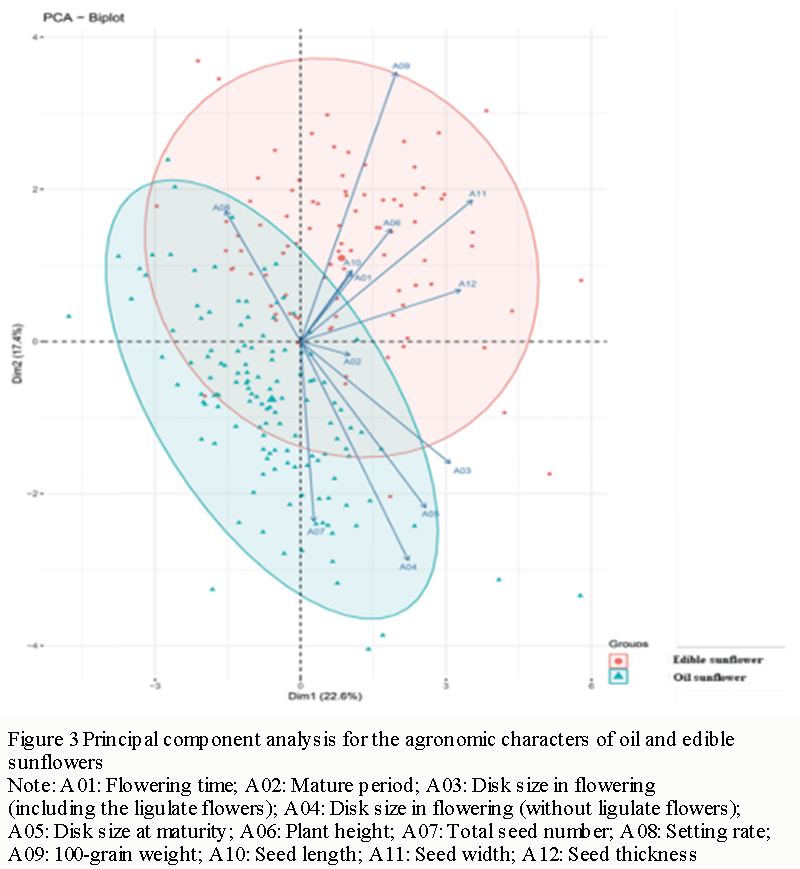 Figure 3 QTL detection of LTG in rice |
 Table 1 QTL affecting LTG of rice detected in RIL population |
1.3 Analysis of candidate gene region
The physical position of the markers on both sides of the QTLs detected in this study was obtained in Gramene (https://archive.gramene.org/). The qLTG-2 corresponded to a physical interval of 0.76 Mb ranged from 9.43 to 10.19 Mb on chromosome 2. A total 108 ORFs predicted were found in the candidate region. The qLTG-4 located in the range of 4.9 Mb between 28.97~33.87 Mb on chromosome 4, and qLTG-7 located in the range of 3.83 Mb on chromosome 7 between 8.88~12.71 Mb. The candidate regions of the qLTG-4 and qLTG-7 were large, and needed to be narrowed by fine mapping.
2 Discussion
LTG tolerance, low-oxygen germination tolerance, and strong germination energy were the basic requirements of DSR varieties (Jin et al., 2001; Jing et al., 2008), which could ensure that DSR overcame the developmental delay caused by low-temperature and guaranteed the vigorous growth of seedling, and is an effective way to solve the problem of poor seedling establishment of DSR. Screening of LTG tolerant resources was the premise of breeding LTG tolerant cultivars. Many important agronomic traits including LTG were complex quantitative traits controlled by multiple genes. Because those traits had low heritability, direct selection for them was difficult. Mapping, isolation, and functional analysis of novel QTLs related to LTG would help to explain the genetic basis of complex agronomic traits, which not only laid the theoretical foundation for selection or improvement of these traits in breeding, but also provided molecular markers for multi-gene polymerization breeding. The most useful principle of QTL mapping was to identify QTL by analyzing genetic markers co-segregated with target traits in segregated populations. In this study, total three QTLs related to LTG were identified by the RIL population from Zhaxima×Nanjing46, which located on chromosome 2, 4 and 7, respectively. The qLTG-2 ranged from 9.43 Mb to 10.19 Mb on chromosome 2, which contained a total 108 predicted ORFs. Interestingly, there was no reported QTL involved in LTG in the candidate region of qLTG-2, which exhibited that the qLTG-2 was a novel locus. The qLTG-4 was located in the range of 28.97~33.87 Mb on chromosome 4, while another QTL related to LTG from USSR5 was mapped in the range of 30.77~34.53 Mb on chromosome 4 by Jiang et al. (2006). The overlapping area of the candidate region of the two QTL was 3.1 Mb, and about 480 predicted ORFs were found in the overlapping area. Since candidate regions of the two QTL had large overlapping region and a lot of genes were contained in the overlapping region, their allelism needed to be further determined. Gong et al. (2009) reported a QTL for LTG on chromosome 7 in “Koshihikari”, which were located in the region of 5.85~12.72 Mb. The qLTG-7 (at 8.88~12.71 Mb) found in this study was also in the candidate region. The common candidate regions of the two QTL were 3.83 Mb, which contained a large number of candidate genes. Therefore, it is necessary to construct secondary segregated populations to fine map them or to carry out allelic tests to further determine whether they were the same allele.
Among the three QTLs identified in this study, the qLTG-2 was a novel QTL locus, which provided a new gen- etic resource for cold-tolerant rice breeding at germination stage. Of these three QTLs, qLTG-7 had the largest effect on PV, which could explain 22.93% of the PV. More importantly, the qLTG-7 was derived from excellent taste quality of japonica rice variety in Jiangsu, which had excellent comprehensive characteristics, and which had higher breeding value. This study laid a good foundation for the future cloning of genes for LTG, and also broadened the material basis for improving LTG of rice, and provided molecular markers for screening LTG.
3 Materials and Methods
3.1 Plant materials
The indica rice varieties “Zhaxima” in Yunnan and the elite japonica rice “Nanjing 46” in Jiangsu were used as parents to establish a RIL population with 135 lines via single seed descent (Fang et al., 2017).
The parents and the RIL population were grown in the experimental field of Jiangsu Academy of Agricultural Sciences in Nanjing at the natural growing season of 2019. The individual spacing and row spacing of rice planting were 13.3 cm×30.0 cm. Field management of the materials in this study was the same as general field of rice.
3.2 Evaluation of seed LTG
Seeds of the parents and the RIL population were dried at 50°C for 7 days to break seed dormancy. The method described by Jiang et al. (2006) and Ji et al. (2008) was used for the evaluation of LTG. The protocol was as follows: One hundred healthy and full seeds of each plant were randomly selected, and surface sterilized for 30 min with 84 Disinfectant Liquid, washed 5 times with sterilized water, and placed on 9 cm diameter petri dishes with two layers of filter paper. And then, 10 mL sterilized water was applied to every dish, and held at 15°C. Germination rate was assessed after 10 days. Seedlings with a coleoptile or radicle longer than 1 mm were taken as germinated. At the end of this test, the seeds were removed to 30℃, and the germination rate was measured again after a further 4 days. Seed were considered free of secondary dormancy if the germination rate was over 80%. All experiments were replicated three times.
3.3 QTL mapping
The linkage map of Zaxima/Nanjing46 RIL population for QTL analysis was composed of 202 SSR markers, evenly distributing on 12 chromosomes (Fang et al., 2017; Zhang et al., 2018). QTL analysis was carried out by QTL IciMapping4.0 (Meng et al., 2015). The LOD threshold of 2.5 was used to declare putative QTLs. The QTL naming was based on the method of McCouch et al. (1997).
QTL physical location and candidate gene analysis were performed in Gramene (https://archive.gramene.org/) and Rice Genome Annotation Project (http://rice.plantbiology.msu.edu/index.shtmL), respectively.
Authors’ contributions
Fang Xianwen and Zhang Suobing were responsible for the experimental design, data analysis, and manuscript writing and revising in this study. Zhang Yunhui, Lin Jing, Chen Haiyuan, Wang Yingjie, Zhu Xiaomei and Song Chunfeng participated in part of the experimental data collection. All authors read and approved the final manuscript.
Acknowledgements
This study was supported by Identification, Evaluation and Innovative Utilization of Quality Special Germplasm Resources of Major Grain Crops (CX (18)1001) and Special subsidy for Chinese medicine public health service in 2018 “National Survey of Chinese Medicine Resources” (2018-43).
Cao W., Wang Y., Tan B., Liu W., Chu L., Pan Z.Y., Zhao G.M., Cao G.Y, and Zhou Y.L., 2018, Evaluation of seed germination vigor of rice cultivars under low temperature or hypoxic condition related with direct seeding rice, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 16(10): 3259-3268
Chen L., Lou Q.J., Sun Z.X., Xing Y.Z., Yu X.Q., and Luo L.J., 2006, QTL mapping of low temperature germinability in rice, Zhongguo Shuidao Kexue (Chinese Journal of Rice Science), 20(2): 159-164
Fang X.W., Zhang Y.H., Xiao X.L., Zhang S.B., Lin J., and Wang Y.J., 2017, Mapping of QTLs for grain shape using recombinant inbred lines in rice (Oryza sativa L.), Jiangsu Nongye Xuebao (Jiangsu Journal of Agricultural Sciences), 33(2): 241-247
Farooq M., Siddique K. H.M., Rehman H., Aziz T., Lee D.J., and Wahid A., 2011, Rice direct seeding: Experiences, challenges and opportunities, Soil & Tillage Research, 111(2): 87-98
https://doi.org/10.1016/j.still.2010.10.008
Fujino K., Sekiguchi H., Sato T., Kiuchi H., Nonoue Y., Takeuchi Y., Ando T., Lin S. Y., and Yano M., 2004, Mapping of quantitative trait loci controlling low-temperature germinability in rice (Oryza sativa L.), Theoretical and Applied Genetics, 108(5): 794-799
https://doi.org/10.1007/s00122-003-1509-4
PMid:14624339
Gong Y.J., Ruan W.J., Xun X., Dong Y.J., Lin D.Z., Ye S.H., and Zhang X.M., 2009, QTL Analysis of cold tolerance for two bud traits in rice, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 7(2): 273-278
Ji S.L., Jiang L., Wang Y.H., Liu S.J., Liu X., Zhai H.Q., Yoshimura A., and Wan J.M., 2008, QTL and epistasis for low temperature germinability in rice, Zuowu Xuebao (Acta Agronomica Sinica), 34(4): 551-556
https://doi.org/10.1016/S1875-2780(08)60021-8
Ji S.L., Jiang L., Wang Y.H., Zhang W.W., Liu X., Liu S.J., Chen L.M., Zhai H.Q., and, Wan J.M., 2009, Quantitative trait loci mapping and stability for low temperature germination ability of rice, Plant Breeding, 128(4): 387-392
https://doi.org/10.1111/j.1439-0523.2008.01533.x
Jiang L., Liu S., Hou M., Tang J., Chen L., Zhai H., and Wan J., 2006, Analysis of QTLs for seed low temperature germinability and anoxia germinability in rice (Oryza sativa L.), Field Crops Research, 98(1): 68-75
https://doi.org/10.1016/j.fcr.2005.12.015
Jin Q.Y., Ou Yang Y.N., Lu Y.L., and Xu Y.C., 2001, Research on some problems of direct-seeded rice in south china and its technical countermeasures, Zhongguo Nongxue Tongbao (Chinese Agricultural Science Bulletin), 17(5): 44-48
Jing D.D., Yu B., Qian H.F., Sheng S.L., Lin T.Z., Hu C.M., Zhang J.B., Diao L.P., Gong H.B., Zhou Y.W., Li C., 2008, Screening and breeding strategies of direct-seeded rice varieties, Jiangsu Nongye Kexue (Jiangsu Agricultural Sciences), (4): 24-27
McCouch S.R., Cho Y.G., Yano M., Paul E., Blinstrub M., Morishima H., and Kinoshita T., 1997, Suggestions for QTL nomenclature for rice, Rice Genetics Newsletter, 14: 11-13
Meng L., Li H., Zhang L., and Wang J., 2015, QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations, The Crop Journal, 3(3): 269-283
https://doi.org/10.1016/j.cj.2015.01.001
Najeeb S., Ali J., Mahender A., Pang Y L., Zilhas J., Murugaiyan V., Vemireddy Lakshminarayana R., and Li Z., 2020, Identification of main-effect quantitative trait loci (QTLs) for low-temperature stress tolerance germination and early seedling vigor-related traits in rice (Oryza sativa L.), Molecular Breeding, 40(1): 10
https://doi.org/10.1007/s11032-019-1090-4
PMid:31975784 PMCid:PMC6944268
Olajumoke B., Juraimi A.S., Uddin M.K., Husni M.H.A., and Alam M.A., 2016, Competitive ability of cultivated rice against weedy rice biotypes-A review, Chilean journal of agricultural research, 76(2): 243-252
https://doi.org/10.4067/S0718-58392016000200015
Schlappi M.R., Jackson A.K., Eizenga G.C., Wang A., Chu C., Shi Y., Shimoyama N., and Boykin D.L., 2017, Assessment of five chilling tolerance traits and GWAS mapping in rice using the USDA mini-core collection, Frontiers in plant science, 8: 1-13
https://doi.org/10.3389/fpls.2017.00957
PMid:28642772 PMCid:PMC5463297
Singh S., Sharma S.N., and Prasad R., 2001, The effect of seeding and tillage methods on productivity of rice-wheat cropping system, Soil & Tillage Research, 61(3-4): 125-131
https://doi.org/10.1016/S0167-1987(00)00188-4
Zhang Y.H., Zhang S.B., Lin J., Wang Y.J., and Fang X.W., 2018, Mining and mapping of QTLs for panicle length from a long-panicle type landrace rice (Oryza sativa L.), Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 16(12): 3967-3972



. PDF(565KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. Suobing Zhang
. Yunhui Zhang
. Jing Lin
. Haiyuan Chen
. Yingjie Wang
. Xiaomei Zhu
. Chunfeng Song
. Xianwen Fang
Related articles
. DSR
. RIL
. LTG
. QTL mapping
Tools
. Email to a friend
. Post a comment